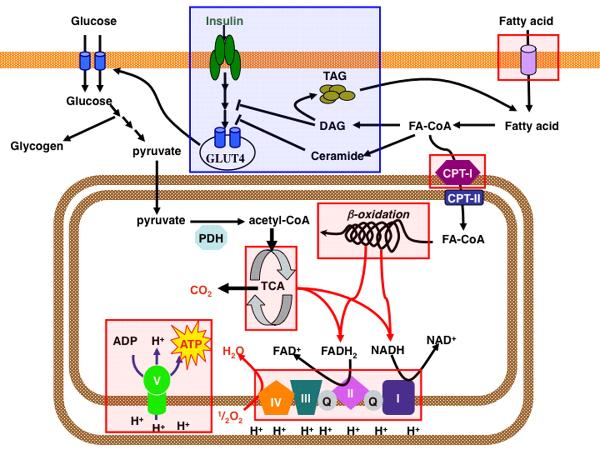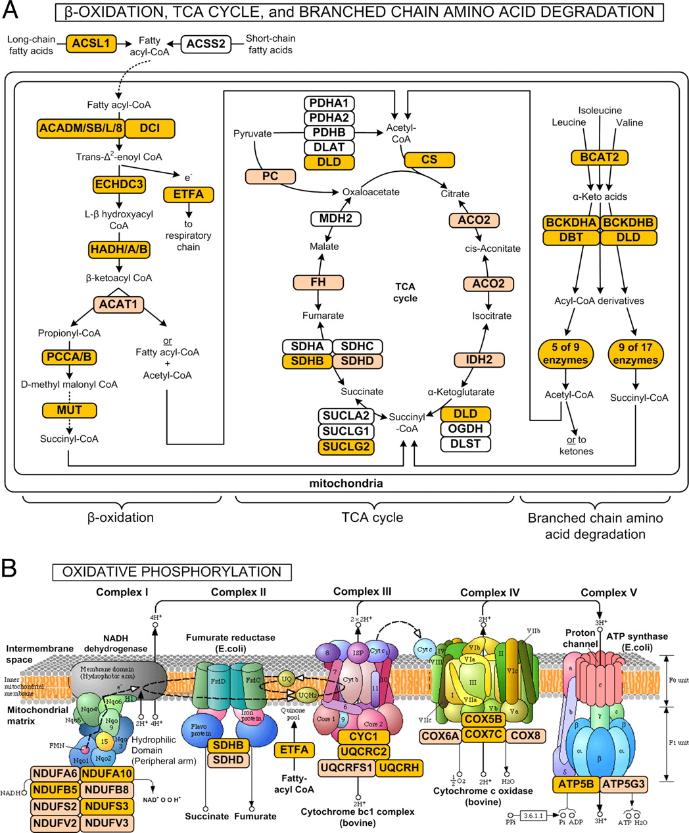
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
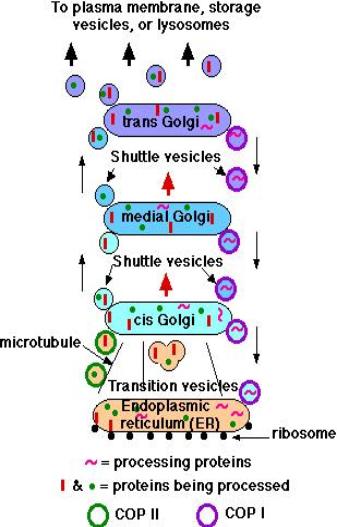 |
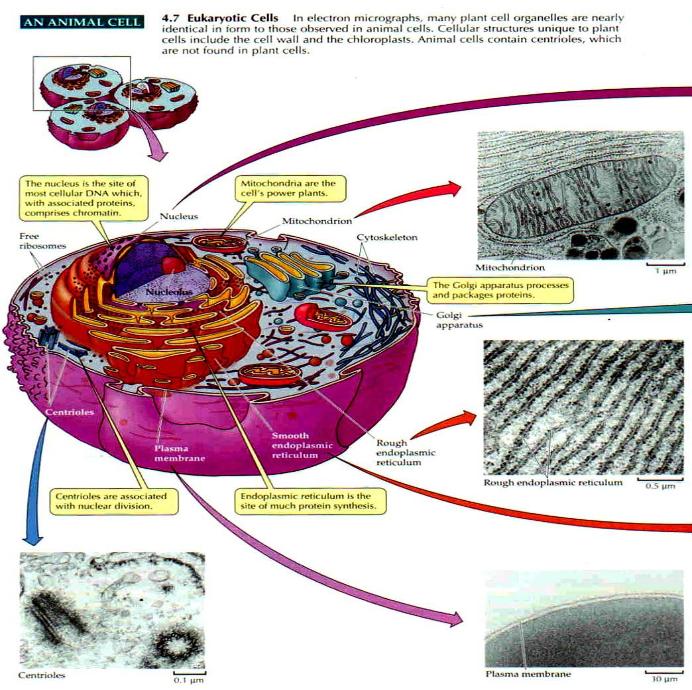
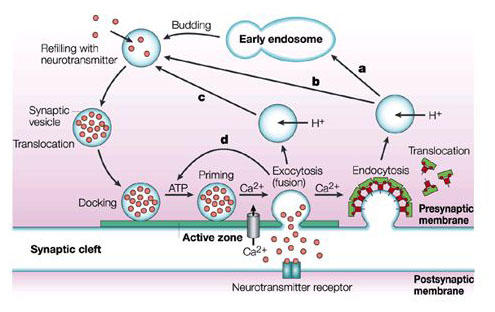
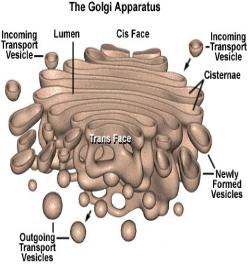
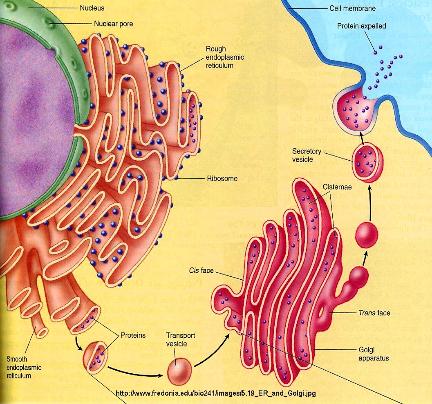
• Transition vesicles pinch off from the surface of the ER carrying integral membrane proteins, soluble proteins awaiting processing, and enzymes • Pinching off requires that the vesicle be coated with COPII (Coat Protein II) • The transition vesicles move toward the cis Golgi on microtubules • As they do so, their COPII coat is removed and they may fuse together forming larger vesicles • These fuse with the cis Golgi • Sugars are added to proteins in small packets so many glycoproteins have to undergo a large number of sequential steps of glycosylation, each requiring its own enzymes • These steps take place as shuttle vesicles carry the proteins from cis to medial to the trans Golgi compartments • At the outer face of the trans Golgi, vesicles pinch off and carry their completed products to various destinations • The movement of cisternal contents through the stack means that essential processing enzymes are also moving away from their proper site of action • Using a variety of signals, the Golgi separates the products from the processing enzymes that made them and returns the enzymes back to the endoplasmic reticulum • This transport is also done by pinching off vesicles, but the inbound vesicles are coated with COPI (Coat Protein I) |
 |
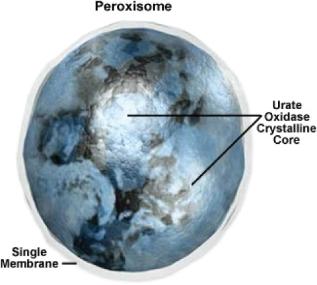 |
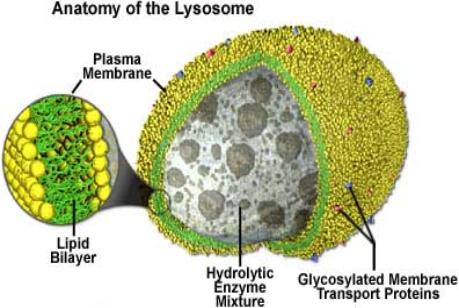
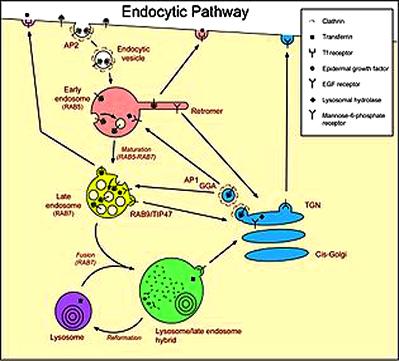 |
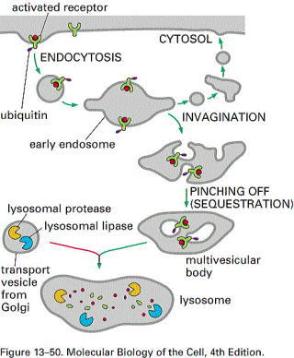 |
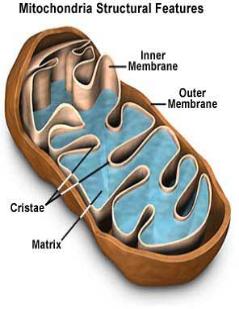 |
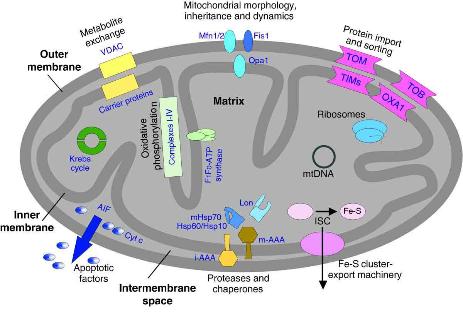 |
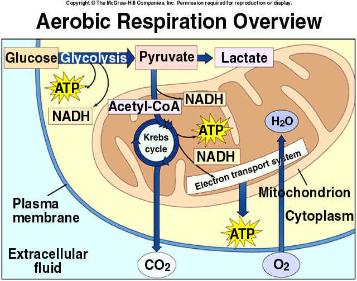 |
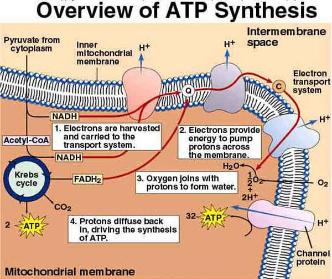 |
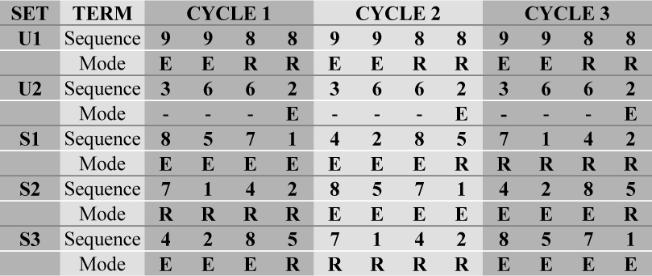
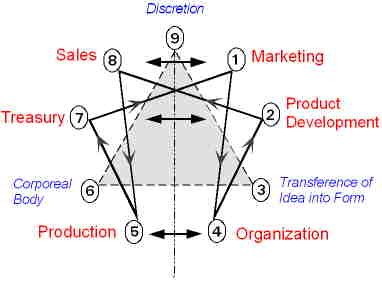 |
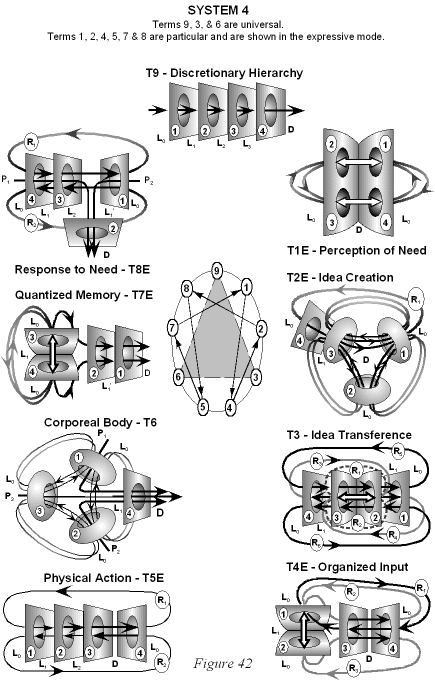
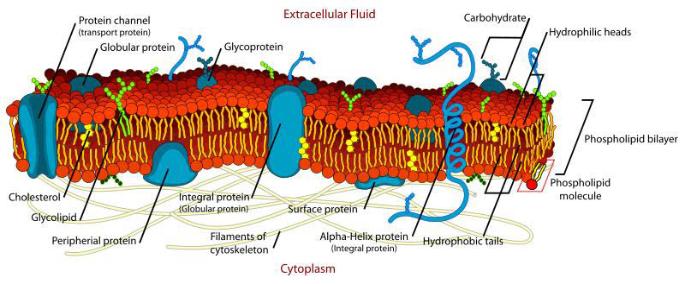
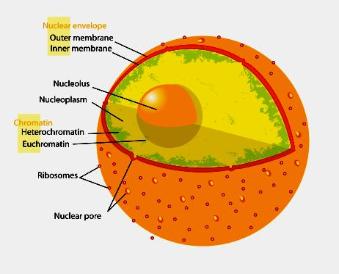
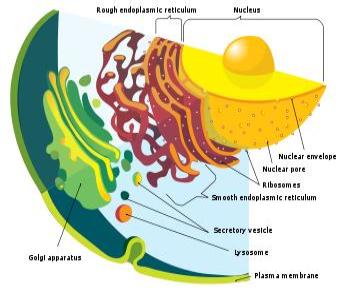
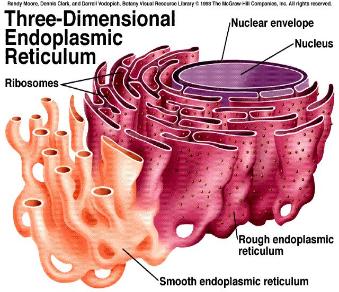
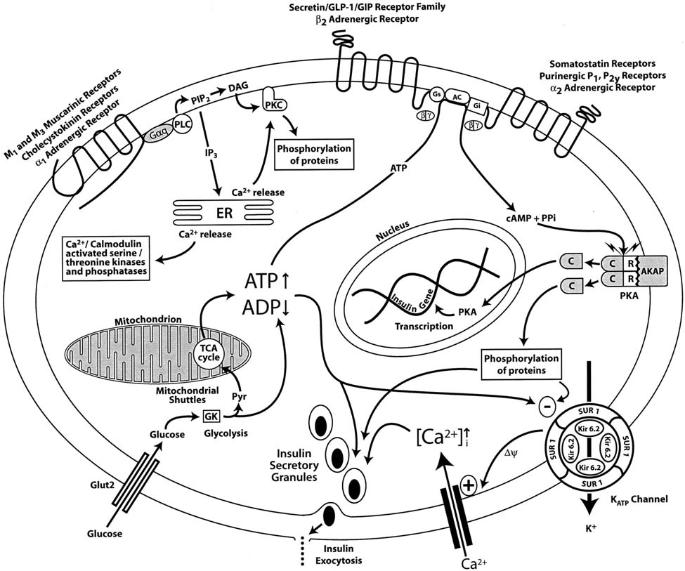
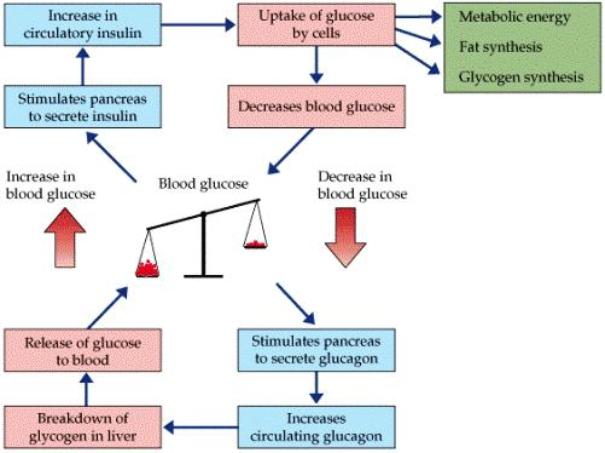
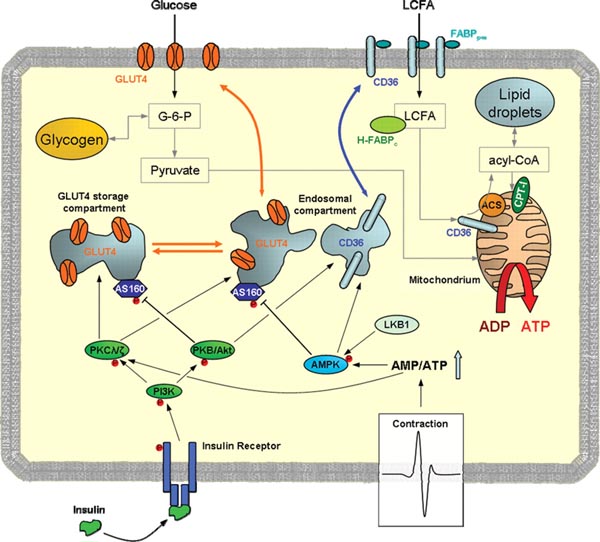
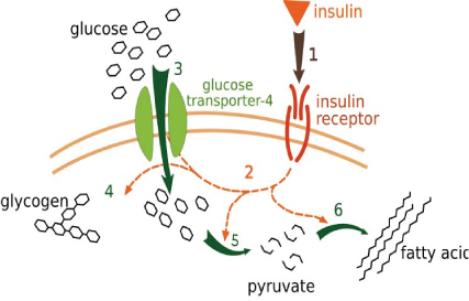
The Particular Terms of System 4 are shown in red as they apply to a business corporation. The flow of work transformation is shown by the connecting lines with arrows. The sequence transformations of the blue Universal Terms are not shown. The polar insights into the Performance, Potential, and Commitment dimensions of the company are shown by the three horizontal arrows respectively. The analogy is useful to understand cell organization and function. |