| Cycle 3 |
T1E |
In Step 4 of this previous cycle, there is an initial assessment of the preparedness of hair cell and muscle action potentials and tone. This can affect the selection of the pattern of cell activation that follows in the next Cycle 1 below. |
| Cycle 1 |
T4E |
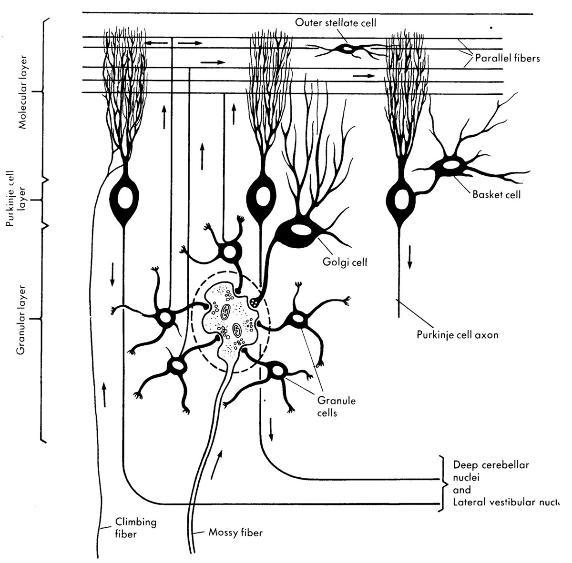
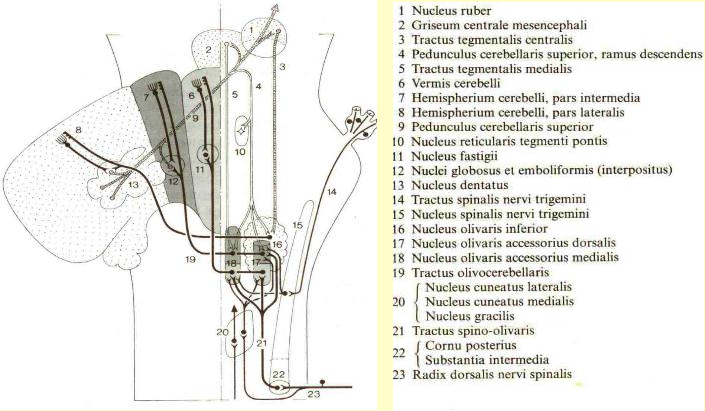
Hair cells activate primary projections of the vestibular system. Muscle spindles monitor parent muscle action and project to relay nuclei in the Central Nervous System (CNS). |
| T2E |
Primary neurons project from hair cells to vestibular nuclei as Idea Terms. Proprioceptive relay nuclei in the CNS also project as Idea Terms to the vestibular nuclei from all spinal levels. |
| T8E |
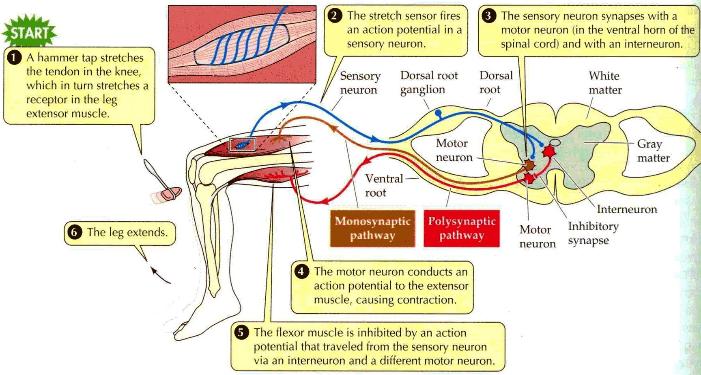
Parallel patterned response outputs that balance sensory vestibular inputs project to eye, neck, and spinal motor nuclei. |
| T5R |
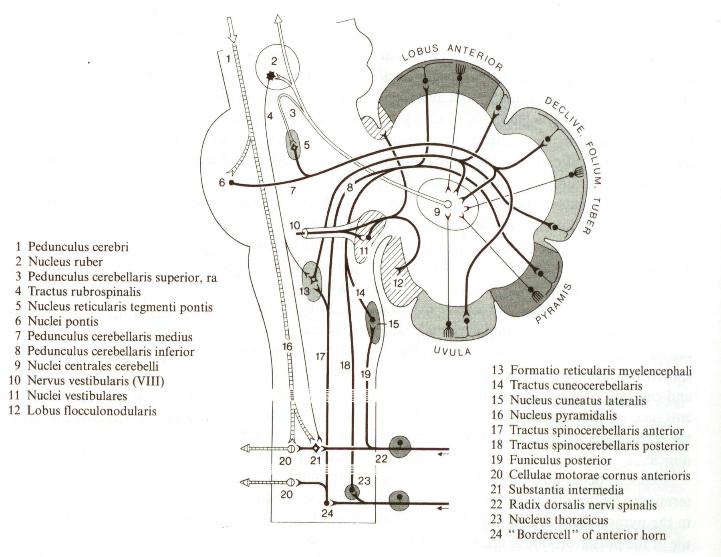
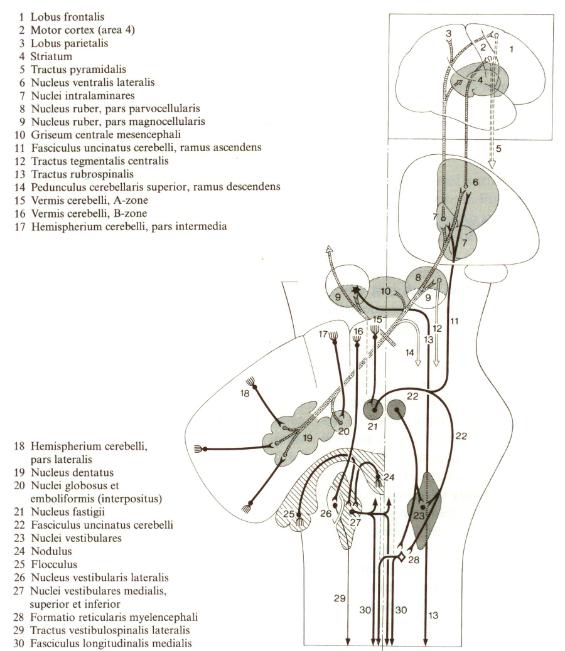
The motor output to eye and neck muscles is reconciled with other inputs to these motor nuclei. The eyes may move to compensate for head movement or move separately. This output is regenerative since it is governed by connections established through prior learning of the individual and the species. The motor output to the neck and eyes is via the Medial Longitudinal Fasciculus (MLF). The motor output to the ventral motor horns of the spinal column is via the lateral vestibulospinal tract. |
| Cycle 2 |
T7R |
Memory of the sequence is stored in dendritic and synaptic protein synthesis of cells involved, as well as the Void. The synaptic connections evolve consistent with learning experience. At the same time related regenerative memories are spontaneously recalled in the reticular formation. This activates efferent projections to hair cells and muscle spindles. |
| T1R |
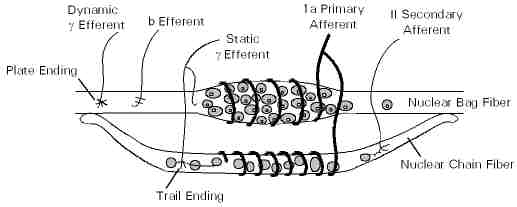
The regenerative efferent projections to hair cells and muscle spindles initiate a patterned motor simulation. In the projection to muscle spindles this takes place in gamma motor neurons that project to the spindles. |
| T4R |
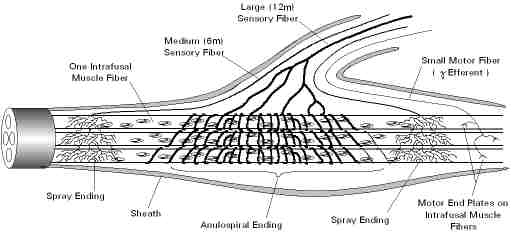
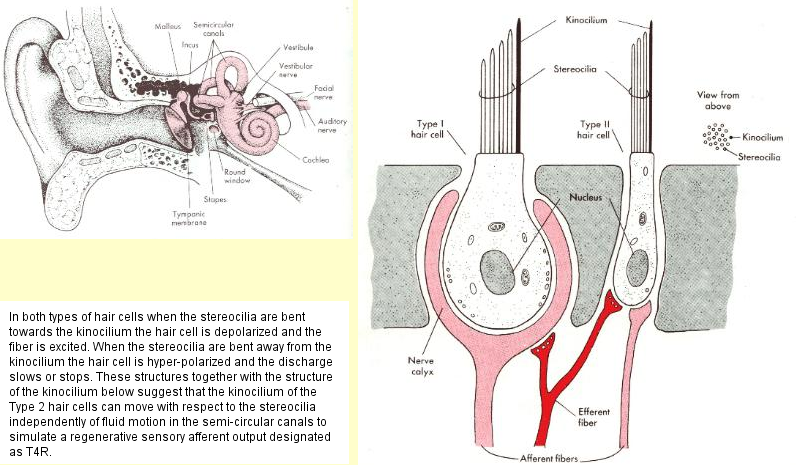
An actual simulation takes place in hair cells and in neck and body muscle spindles. The hair cell simulation involves the active movement of the kinocilium in the Type 2 hair cell with respect to the current state of the Type 1 hair cells. The muscle spindle simulation likewise simulates a pattern of movement with respect to the current position of muscles. |
| T2R |
The simulations project as regenerative vestibular input and likewise corrective proprioceptive feedback to vestibular nuclei as a regenerative idea to restore balance. |
| Cycle 3 |
T8E |
The simulations result in a synapse in the vestibular nuclei to a pattern of planned motor output to eye, neck and body muscles via the MLF and the lateral vestibulospinal tracts as in the T8E Term above. All Particular Set T8 Terms are expressive. (See Part 1). |
| T5E |
The output of vestibular nuclei to eye, neck and body spinal motor nuclei for corrective action is reconciled with other inputs to these nuclei as they project to muscles. This output is expressive since it is creatively planned by proprioceptive input. It is patterned action of the Host (1) that directs Organs (2) whereas in the regenerative sequence it is conditioned Organ (2) processes established through experience that direct the Host (1). (See Part 1) |
| T7E |
Memory of the action sequence is stored and expressive memories recalled similar to T7R above. |
| T1E |
There is a perception of the capacity to respond to ongoing vestibular needs. This involves the recovery of cells processes such as action potentials and the stabilization of fluid in semi-circular canals, preparatory for the next action sequence. |